
Research Professor
Department of Neuroscience
1971-74 B.Sc. Hebrew University (Biology)
1975-81 Ph.D. Cornell University
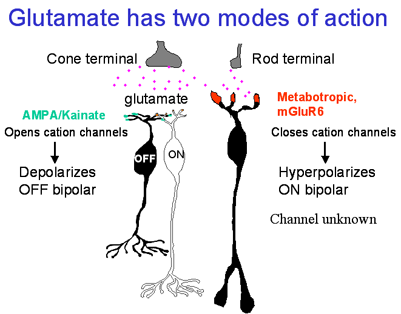
Schematic of information transfer at the first visual synapse. Rod photoreceptors contact a single type of second order neuron, the rod bipolar cell. Cone photoreceptors contact 10 types of second order neurons of two classes: ON and the OFF cone bipolar cells. Glutamate has two modes of action: by the conventional AMPA/kainate receptor, it depolarizes the OFF bipolar cells; by the metabotropic glutamate receptor mGluR6, it hyperpolarizes ON bipolar cells.
The retina provides an excellent model system for signal processing because the input (visual image) is well defined, the output (ganglion cell responses) has been thoroughly studied, and circuits of parallel pathways from photoreceptor to ganglion cells have been described better than any other neural system. To reliably transfer the signal from photoreceptor to ganglion cells through these pathways under a large range of luminances, the retina employs gain control and noise removal mechanisms. Gain is adjusted by GABAergic and glycinergic feedback circuits in the outer and inner plexiform layers. These circuits tune photoreceptor, bipolar and ganglion cells' responses by averaging and feeding back ambient light information.
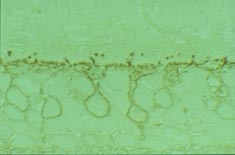
Staining for the alpha subunit of Go is localized to somas and dendrites of rod bipolar and ON cone bipolar cells.
A major focus of the lab is the ON bipolar cell and its signaling cascade. The ON bipolar cell expresses the metabotropic glutamate receptor mGluR6. This receptor activates the G protein Go which, through an intermediary cascade of proteins, closes a nonspecific cation channel. We are studying the events that occur downstream of Go and how they are modulated. Whether Go activates the channel directly, or through a second messenger is important because the manner of activation determines the gain and the adaptation of these cells. In particular, both mGluR6 and Go are present in rod and cone bipolar cells, but the two cells have responses with different kinetics — slow for night vision vs. fast for day vision — so one would expect different cascades. In addition, we study how the proteins in the cascade complex are trafficked to the dendritic tip, and how a lack of a certain element destabilizes the complex. Knowledge of this cascade is essential for understanding and treating night blindness. A second project concerns cone bipolar cell types; there exist about 5 types that are thought to conduct different temporal bandwidth. Nothing is known about their precise light properties and how they achieve their differences. We study this by calcium imaging of ON bipolar cells. Such understanding will shed light on daylight vision.
Electrophysiology, immunocytochemistry, electroretinogram, dye injection, electron microscopy, molecular biology, yeast two hybrid systems, computer simulation, live two photon imaging